高山克荒,大川利涉:从血管正常化的重要性聊聊康方生物AK112在免疫治疗的优势(万字长文)



一些朋友问我,康方生物AK112的机制,是否有可行性,康方生物AK112在已经确定是更好的抗血管药物(安全性太明显)的前提下,是否也是更好的免疫治疗药物。正好,最近有一篇名为Linking tumour angiogenesis and tumour immunity(Kabir AU, Subramanian M, Kwon Y, Choi K. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z)的综述,我觉得写得很好了,如果有能力,我推荐看原文,一定会有所收获的,我在这里结合自己的想法和观点适当的删增,分享给大家,和大家交流一下讨论下。必须承认的是,文章肯定是有难度的,我也会尽量标出重点总结结论,希望可以和大家多多交流多多讨论,也希望我们都能有所收获,实在看不懂直接看最后的结论也是可以的,结论应该可以看懂。
免疫检查点阻断疗法(Immune checkpoint blockade)彻底改变了转移性和实体瘤的治疗,在一部分的患者中实现了持久的反应。然而,大多数患者对免疫检查点阻断没有反应,这突出表明迫切需要更好地理解治疗效力的决定因素。有效抗肿瘤免疫反应的一个关键障碍是肿瘤相关血管的异常结构和功能,这阻碍了免疫细胞浸润并有助于免疫抑制性肿瘤微环境的发展。当前的研究发现了肿瘤微环境内血管生成与免疫活性之间的负相关。
首先,肿瘤细胞固有的异质性是导致癌症患者治疗抵抗的一个主要因素,与在患者和肿瘤类型之间差异很大的遗传不稳定和异质性的癌细胞相比,肿瘤微环境(tumour microenvironment, TME)的许多特征在不同癌症中相对稳定且保守。肿瘤微环境的这些稳定特征使其成为治疗干预的一个有吸引力的靶点,特别是考虑到其在促进免疫逃逸和对各种治疗(包括免疫疗法和抗血管生成药物)抵抗中的关键作用。因此,针对 TME 的策略可能对多种肿瘤类型广泛有效。
那么,肿瘤微环境TME 作为一个复杂的细胞龛,控制着肿瘤的生长和转移。它包含各种免疫细胞、基质细胞(如内皮细胞和成纤维细胞)以及细胞外基质成分。近期的临床前证据强调了肿瘤内血管功能障碍的关键影响。肿瘤血管本质上是迂曲和渗漏的,面临着不断扩大的肿瘤块带来的日益增加的机械压力,导致不连续的血管系统和可变的灌注。这种血管异常阻碍了治疗药物的递送。此外,增殖的肿瘤细胞消耗氧气,加上它们产生的代谢废物和氧气输送不佳,创造了一个酸性和缺氧的微环境,加剧了血管功能障碍。肿瘤血管系统的高通透性和不成熟特性限制了细胞毒性 T 细胞的有效浸润及其对肿瘤细胞的靶向作用。低灌注相关的糖酵解适应进一步促成了一个损害 T 细胞效应功能的酸性环境。而且,肿瘤内皮细胞可以表达死亡信号,如 FAS 配体(FAS ligand , FASL),诱导细胞毒性 T 细胞凋亡。新兴的研究强调了肿瘤血管生成与浸润免疫细胞之间的相互反馈回路。抑制肿瘤血管生成可增强肿瘤免疫,而肿瘤免疫反过来又促进血管正常化。相反,免疫抑制细胞刺激血管生成,这进一步增强其他免疫抑制细胞的招募,维持肿瘤生长和转移。
肿瘤血管的特征
血管生成开关 (The angiogenic switch)
血管生成是一个涉及多种细胞类型、细胞外基质成分和一系列促血管生成因子的复杂过程。它可以分为不同的阶段:(1) 蛋白酶降解现有血管的基底膜,(2) 内皮细胞活化与迁移,(3) 这些内皮细胞的一个亚群的增殖,以及 (4) 形成与现有血管网络相连的管腔和管状结构。在成人中,血管生成受到促血管生成和抗血管生成因子之间平衡的严格调控,通常发生在女性生殖周期、组织再生和伤口愈合期间。
在早期的微观肿瘤集落中,营养物质和废物的交换主要通过简单扩散进行。然而,当肿瘤块生长到大约 2–3 mm 大小时,仅靠扩散变得不足以满足增加的代谢需求。为了应对肿瘤核心的缺氧,缺氧诱导因子(HIF)诱导血管内皮生长因子(VEGF)的表达,刺激附近血管网络形成毛细血管芽,确保持续的营养和氧气供应,同时能够从不断扩大的肿瘤中清除代谢废物和二氧化碳。这一关键转变被称为“血管生成开关”。没有这个开关,肿瘤集落面临坏死或凋亡的风险,并可能无限期保持休眠状态。
与生理性血管生成(其中血管形成受到严格调控并导致功能成熟)不同,肿瘤细胞和 TME 内的免疫抑制细胞持续产生促血管生成因子,驱动持续的血管生成刺激。过度和持续的血管生成刺激导致异常血管形成,其特征是不成熟、扩张和结构缺陷的血管,通常缺乏适当的周细胞覆盖,最终导致功能失调的血管。这些异常的、无功能的血管无法有效输送氧气和营养物质,创造了一个缺氧和免疫抑制的 TME,这进一步促进了肿瘤进展和治疗抵抗。它们增加的通透性通过促进肿瘤细胞进入循环进一步促进转移。在继发部位,微转移灶与原发。在 20世纪70年代初期,Judah Folkman开创了肿瘤血管生成对于实体瘤生长至关重要的概念,并提出抑制血管生成作为一种潜在的治疗方法。从那时起,许多血管生成调节因子被识别出来,抗血管生成治疗已成为一种重要的癌症治疗策略。尽管有前景,但抑制血管生长常常导致显著的副作用,并且仅针对血管生成已被证明不足以完全消除肿瘤生长。这部分是由于治疗药物的功能欠佳以及错误判断了抗血管生成药物可能最有效的治疗背景,也因此给康方生物AK112留下了空间。当芽生血管生成被阻断时,肿瘤也可以通过激活代偿途径来适应,导致替代形式的血管生成。这些替代方式包括血管套叠(预先存在的血管腔分裂形成新的分支)、血管共择(肿瘤细胞沿着预先存在的血管排列)和血管拟态(肿瘤细胞模仿内皮细胞形成促进血流的管状结构)。
肿瘤血管生成因子以外的血管生成调节因子 (Angiogenic regulators beyond tumour angiogenesis)
促血管生成调节因子包括一系列刺激血管形成以支持组织生长和存活的生长因子、酶和信号分子。关键的促血管生成因子包括 VEGF、成纤维细胞生长因子 2(FGF2,也称为碱性成纤维细胞生长因子(bFGF))、血小板衍生生长因子(PDGF)、转化生长因子-α(TGFα)和 TGFβ、表皮生长因子(EGF)、肝细胞生长因子(HGF)、胎盘生长因子(PIGF)、基质金属蛋白酶(MMPs)、组胺、1-磷酸鞘氨醇(S1P)、IL-8、apelin(APLN)和血管生成素 I(ANGI)及 ANG2。这些因子,连同代谢和机械应激源,如缺氧、营养缺乏、低 pH 值和活性氧物种,促进肿瘤血管生成。相反,一系列内源性血管生成抑制剂起到平衡这些促血管生成信号的作用。这些包括血管抑素(angiotastin)、内皮抑素(endostatin)、arrestin、canstatin、tumstatin、IL-12、血小板反应蛋白 I(TSPI)和 TSP2、金属蛋白酶组织抑制剂(TIMPs)以及干扰素-α(IFNα)、IFNβ 和 IFNγ。重要的是要注意,这些血管生成因子存在许多细微差别和功能双重性。例如,VEGF-A 是所有条件下的原型血管生成因子,而 VEGF-B 是内皮细胞的关键存活因子,并被报道抑制病理性血管生成。TGFβ 在早期阶段通过抑制细胞周期进程和诱导凋亡而作为肿瘤抑制因子,但在后期通过支持血管生成和诱导上皮-间质转化来促进肿瘤进展。ANGI 促进血管成熟和新形成血管的稳定,但 ANG2 诱导血管失稳、周细胞丢失、出芽和血管生成。尽管许多研究集中在利用血管生成因子来抑制肿瘤血管生成并有效地“饿死”肿瘤,但许多促血管生成生长因子对于维持血管稳态和功能是必需的。因此,治疗性抑制关键的促血管生成因子可能导致不良副作用,包括严重的并发症,如咯血和内穿孔,这又给康方生物AK112留下了空间。此外,血管生成因子可以独立地抑制抗肿瘤免疫并促进 TME 内的免疫抑制。由肿瘤细胞、基质细胞和包括肿瘤相关巨噬细胞(TAMs)在内的免疫细胞产生的高水平 VEGF 不仅促进血管生成,而且还促进髓源性抑制细胞(MDSCs)的迁移和分化,招募调节性 T 细胞(Treg细胞)并抑制树突状细胞成熟。PDGF 以多种方式影响树突状细胞,包括上调树突状细胞上的 CLEC2 表达,从而诱导 T reg细胞分化并抑制 CD4+ T 细胞增殖。用 PDGF-BB 亚型处理的树突状细胞也显示出改变的细胞因子产生,即 TNF 减少和 IL-10 产生增加。然而,有趣的是,PDGF-DD亚型作为自然杀伤细胞上 NKp44 受体的配体,刺激它们分泌 TNF 和 IFNγ,从而在胶质母细胞瘤中抑制肿瘤生长。此外,在 MC38 肿瘤中,源自 TAMs 的 FGF2 驱动 TAM 重塑朝向促肿瘤表型,并且是头颈鳞状细胞癌细胞和人未成熟树突状细胞中 PDL1 表达的强效诱导剂。FGF 信号的小分子抑制剂可以通过 STAT1 非依赖性途径有效阻断 PDL1 上调并抑制 IFNγ 介导的 PDL1 表达。此外,肿瘤分泌的 FGF21 通过 AKT-mTORC1-固醇调节元件结合蛋白 1(SREBP1)信号通路重编程 CD8+T 细胞中的胆固醇代谢,从而破坏抗肿瘤免疫,导致胆固醇生物合成增加和 T 细胞耗竭。这些发现强调了血管生成因子如何独立地促进免疫抑制和肿瘤进展。
图片来源:Kabir AU, Subramanian M, Kwon Y, Choi K. Linking tumour angiogenesis and tumour immunity. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z
肿瘤血管生成与肿瘤免疫之间的反馈调节 (Feedback regulation between tumour angiogenesis and tumour immunity)
异常肿瘤血管负向调节肿瘤免疫 (Abnormal tumour vessels negatively regulate tumour immunity)
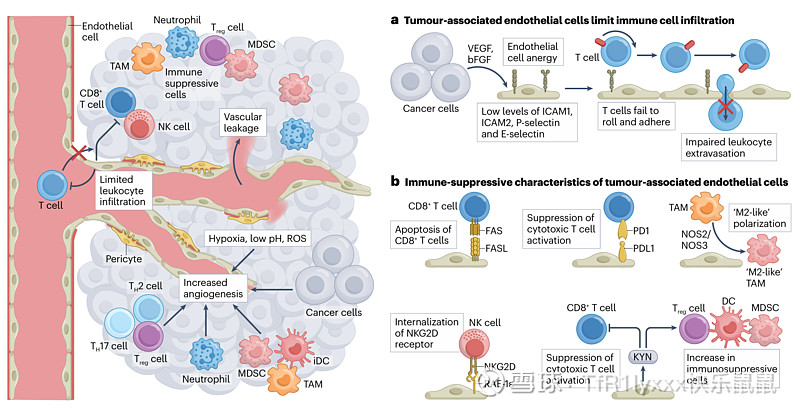
内皮细胞在调节免疫细胞运输中的作用已得到充分证实。免疫细胞从循环系统外渗到组织中主要通过一个称为旁细胞跨内皮迁移(paracellular diapedesis)或跨内皮迁移(transendothelial migration)的过程发生。这个过程通常发生在毛细血管后微静脉,依赖于内皮细胞和免疫细胞双方粘附分子和趋化因子之间的相互作用。跨内皮迁移包括几个步骤:选择素介导的栓系和滚动、整合素依赖性粘附和牢固停滞、管腔内爬行以及最终的跨迁移。在正常生理条件下,免疫细胞迁移主要局限于特化组织,如初级和次级淋巴器官、胸腺和骨髓。在健康组织中,炎症细胞因子如 TNF、IFNγ 和 IL-1β 通过上调粘附分子并重新排列它们以促进最佳免疫细胞通道来启动内皮细胞进行白细胞跨迁移。然而,在 TME 中,肿瘤内皮细胞通常下调关键的粘附分子,如细胞间粘附分子 1(ICAM1)、ICAM2、血管细胞粘附分子 1(VCAM1)和选择素(P-选择素和 E-选择素),这种下调损害了免疫细胞外渗和招募到肿瘤块。虽然上述粘附分子通常被认为在免疫细胞跨内皮迁移中起作用,但不同的粘附分子和趋化因子可能导致免疫亚群的不同招募。例如,当清道夫和粘附受体 stabilin-1(也称为 clever-1)在肿瘤内皮上表达时,它增强了单核细胞、巨噬细胞和Treg细胞向肿瘤的浸润。当凋亡信号调节激酶 1(ASK1)由肿瘤内皮表达时,它促进单核细胞和巨噬细胞的招募,而不影响 T 细胞的招募。此外,肿瘤内皮表达的 CX3C-趋化因子配体 1(CX3C11)优先调节表达 CX3C-趋化因子受体 1(CX3CR1)的单核细胞和巨噬细胞向肿瘤块的浸润。肿瘤内皮细胞中质膜囊泡相关蛋白(PLVAP)的表达促进了表达叶酸受体-β(FOLR2)的巨噬细胞的招募。相比之下,用黑色素瘤分化相关蛋白 5(MDAS)的激动剂刺激内皮,诱导 I 型干扰素和趋化因子(如 CXCL9 和 CXCL10)的释放,这些因子支持效应 T 细胞浸润到肿瘤中。值得注意的是,在 VEGF 和 FGF2 等血管生成因子的影响下,肿瘤内皮细胞对炎症刺激的反应性降低,这种状态称为内皮细胞无反应性(endothelial cell anergy),这进一步损害了白细胞-内皮细胞相互作用,加剧了 TME 的免疫抑制性质
除了限制免疫细胞浸润外,肿瘤内皮细胞还在促进免疫抑制环境中发挥积极作用。它们表达抑制性蛋白,如 PDLL、PDL2、TIM3 和吲哚胺 2,3-双加氧酶(IDO),这些蛋白抑制细胞毒性 T 细胞的活化和功能。此外,肿瘤内皮细胞通过表达凋亡诱导配体(如 FASL 和 TRAIL)对细胞毒性 T 细胞形成选择性屏障。这些配体主要诱导细胞毒性 T 细胞凋亡,而表达抗凋亡蛋白(如 cFLIP)的Treg细胞则得以幸免。肿瘤内皮细胞通过分泌可溶性因子(包括 TGFβ、PGE2、IL-6、IL-10 和 IL-35)进一步促进免疫抑制条件。而且,它们上调 RAF-1e,这是一种配体,可触发自然杀伤细胞上 NKG2D 受体的内化,从而抑制其抗肿瘤活性。此外,肿瘤内皮细胞中一氧化氮合酶 2(NOS2)和 NOS3 的表达可以影响 TAMs 向促肿瘤 M2 样巨噬细胞表型的重塑,加剧免疫抑制。
异常肿瘤血管也可以通过创造缺氧的 TME 来间接抑制肿瘤免疫。组织不良和功能失调的血管阻碍了有效的氧气输送,导致肿瘤内出现缺氧区域。缺氧反过来又促进 MDSCs 的招募和分化,并通过降低 T 细胞受体信号传导的强度来抑制效应 T 细胞的活性。HIF-1α 还促进 CD39 和 CD73 的表达,以增强浆细胞样树突状细胞的浸润,导致 T reg细胞抑制细胞毒性 CD8+T 细胞。HIF-1α 还被显示可招募骨髓来源的 CD45+髓系细胞,这些细胞包含 Tie2+、VEGFR1+、CD11b+和 F4/80+亚群,以及内皮和周细胞祖细胞,以促进胶质母细胞瘤中的新血管形成。在这项研究中,骨髓来源的 CD45+细胞的 MMP9 活性对于通过增加 VEGF 生物利用度来启动血管生成是必需且足够的。此外,在缺氧 TME 中诱导的 HIF-1α 维持 Treg 细胞的适应性。血管异常还导致酸性的 TME,其特征是乳酸和其他酸性代谢物的积累。TME 的低 pH 值诱导浸润的效应 T 细胞出现无反应性。高水平的乳酸抑制细胞毒性 T 细胞释放 IL-2、IFNγ、穿孔素和颗粒酶。此外,乳酸使 TAMs 偏向促肿瘤 M2 型表型,并促进促肿瘤中性粒细胞的浸润。
免疫抑制细胞促进肿瘤血管生成 (Immune-suppressive cells promote tumour angiogenesis)
肿瘤微环境内的免疫反应是由来自先天和适应性免疫系统的髓系和淋巴样细胞之间复杂的相互作用所塑造的。这些反应可以大致分为促肿瘤或抗肿瘤。简而言之,促肿瘤免疫反应通常由 MDSCs、M2 样巨噬细胞、促肿瘤中性粒细胞以及调节性 B 细胞和 Treg细胞介导。相反,抗肿瘤免疫通常以树突状细胞、炎症性(M1 样)巨噬细胞、自然杀伤细胞以及效应 B 细胞和 T 细胞的功能为特征。对肿瘤进展的净影响取决于这些相反免疫过程之间的平衡,这些过程通常在局部和远处肿瘤部位(如骨髓)同时发生。值得注意的是,免疫抑制细胞,如 MDSCs、Treg细胞和 TAMs,通过分泌促血管生成因子(如 VEGF、TGFβ 和 FGF2)来促进肿瘤血管的生成。同时,这些免疫抑制细胞通过释放细胞因子和信号分子(如 IL-10、TGFβ 和精氨酸酶)来抑制抗肿瘤免疫反应,这些分子抑制效应 T 细胞和自然杀伤细胞的活性,进一步使肿瘤能够逃避免疫检测和破坏。这些过程的累积效应是增强血管生成和维持免疫抑制性 TME,这支持肿瘤扩张和对包括免疫疗法在内的治疗的抵抗。
血管正常化改善抗肿瘤免疫 (Vessel normalization improves antitumour immunity)
血管正常化概念 (Vessel normalization concept)
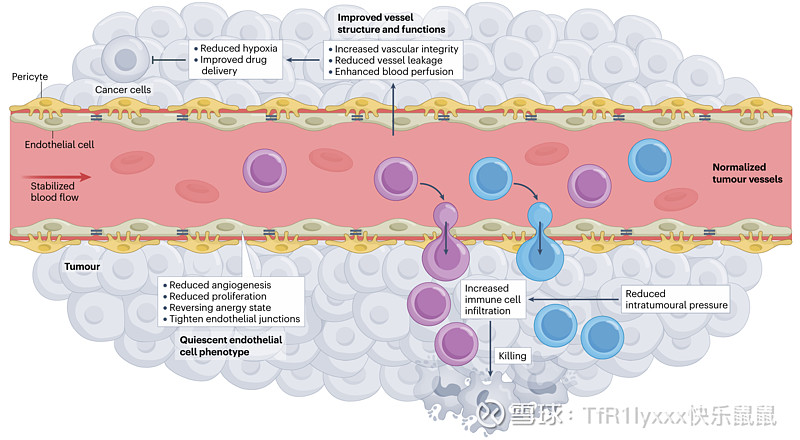
在2001年的一篇开创性论文中,Rakesh Jain提出了“肿瘤血管正常化”的概念,表明纠正肿瘤中促血管生成和抗血管生成功能之间的不平衡可以将异常血管系统恢复到更功能性的状态。与旨在完全根除肿瘤血管的策略不同,血管正常化侧重于改善其结构和功能。这种方法被认为通过增强血液灌注、减少血管通透性和瘤内缺氧,并促进更有效地递送细胞毒性药物来改善肿瘤控制。许多实验室观察到,现有抗血管生成药物的合理治疗方案在不同程度上促进了血管正常化,为与放疗、化疗和免疫疗法的不同联合治疗提供了机会。
图片来源:Kabir AU, Subramanian M, Kwon Y, Choi K. Linking tumour angiogenesis and tumour immunity. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z
血管正常化导致内皮细胞抗原呈递和处理的上调 (Vessel normalization leads to upregulation of antigen presenting and processing in endothelial cells)
正常化的肿瘤内皮细胞,虽然显示出生长因子活性和血管化减少的证据,但通常表现出涉及抗原加工和呈递的通路上调。这引出了一个问题:肿瘤内皮细胞是否能主动呈递抗原,如果是,这在肿瘤背景下可能扮演什么角色?虽然专业的抗原呈递细胞,如树突状细胞、B 细胞和巨噬细胞,对于启动抗肿瘤免疫反应至关重要,但内皮细胞也在其表面表达 MHC 分子和共刺激蛋白。内皮细胞上 MHC 和共刺激分子的表达水平因物种和病理条件而异,但大量研究支持内皮细胞呈递抗原的能力,表明内皮细胞在调节免疫反应中扮演更积极的作用。在肿瘤背景下,淋巴管内皮细胞(LECs)已被显示上调 MHC II 类和相关抗原加工机制,以促进黑色素瘤模型中Treg细胞的扩增。在 LECs 中特异性缺失 MHC II 类的小鼠中,肿瘤生长减少,并且肿瘤内效应 CD4+ T 细胞的浸润增加。在 TME 中 LECs 缺乏 MHC II 类表达的肿瘤中,细胞的抑制功能被深刻改变。此外,LECs 中的 IFNγ 信号增强了它们交叉呈递细胞毒性 T 淋巴细胞(CTL)介导的肿瘤细胞死亡后释放的肿瘤抗原的能力,这导致 CTL 对 LECs 的抗原特异性杀伤。当 LECs 中 IFNγ 受体被删除时,这种杀伤效应被废除,证明 IFNγ 对于减少肿瘤相关淋巴管密度和引流至关重要。在多发性骨髓瘤中,骨髓内皮细胞低水平表达共刺激分子,如 CD40、CD80、CD86 和 ICOSL。它们呈递抗原并刺激表达 FOXP3 并分泌 IL-10 和 TGFβ 的中心记忆 CD8+T 细胞,通过抑制由常规抗原呈递细胞激活的效应 CD8+T 细胞来促进促肿瘤活性。肿瘤血管系统的单细胞转录组分析显示,具有抗原呈递表型的 LECs(apLECs)富含与抗原呈递相关的基因,并显示出与血管生成尖端样 LECs 不同的特征。在肝细胞癌(HCC)患者中,较高比例的 apLECs 与改善的生存相关,与以尖端样 LECs 为主的患者的结果形成对比。
这些发现表明,内皮细胞在特定条件下可以超越其在血管生成中的经典作用,并有助于免疫调节。然而,重要的是要注意,尚未有研究报道肿瘤内皮细胞是否可以呈递肿瘤来源的抗原。此外,尚不清楚这种抗原呈递将如何影响整体肿瘤免疫,这使得这成为未来研究中一个非常有趣的领域。血管生成性和抗原呈递性内皮细胞是否代表不同的群体是另一个有趣的问题,可能为内皮异质性和特化功能提供重要见解。可能地,在经历血管正常化的 Myct1敲除肿瘤内皮中识别出的一个具有抗原呈递和处理通路的“过渡性内皮细胞”亚群,支持了抗原呈递内皮细胞可能代表一个独特功能群体的观点。
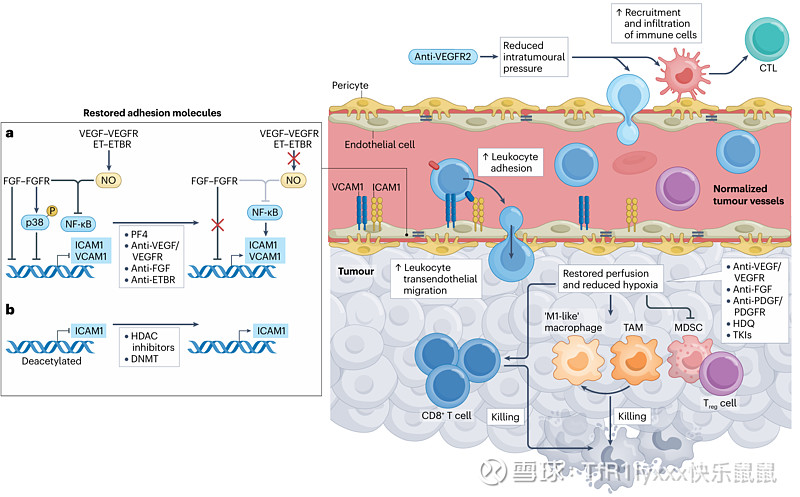
血管正常化增强免疫细胞向肿瘤的跨内皮迁移 (Vessel normalization enhances transendothelial migration of immune cells into tumours)
在过去的30年里,对内皮细胞无反应性更深入的机制理解导致了更有针对性的疗法来对抗这种现象。早期研究通过干扰肿瘤血管生成过程显示了内皮细胞无反应性的逆转。值得注意的是,诸如使用 TKIs、靶向 VEGF 的单克隆抗体和靶向 FGF2 的单克隆抗体阻断血管生成等干预措施都已证明改善了白细胞粘附和跨迁移进入肿瘤组织。血小板因子 4(PF4),一种内源性的肿瘤血管生成抑制剂,被显示可以恢复血管生成因子诱导的 ICAMI 下调,促进肿瘤内皮对炎症刺激的反应性,并改善白细胞与内皮细胞的粘附。在另一项研究中,肿瘤内皮细胞中 ICAMI 的抑制归因于 ICAMI 启动子位点的组蛋白去乙酰化,这可以通过使用组蛋白去乙酰化酶抑制剂和 DNA 甲基转移酶来逆转。ICAMI 表达的恢复与黑色素瘤体内模型中白细胞粘附增加和白细胞向肿瘤核心的浸润改善相关。在其他研究中也观察到了类似的机制驱动的肿瘤内皮细胞无反应性逆转。当高磷酸化的 p38 MAP 激酶(响应 FGF2 刺激而发生)被抑制时,通过 NF-κB 激活恢复 ICAMI 表达和白细胞粘附。一氧化氮抑制也增加了无反应性内皮细胞对炎症细胞因子的敏感性,并且阻断内皮素 B 受体促进 T 细胞与内皮的粘附及其随后的跨迁移。
图片来源:Kabir AU, Subramanian M, Kwon Y, Choi K. Linking tumour angiogenesis and tumour immunity. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z
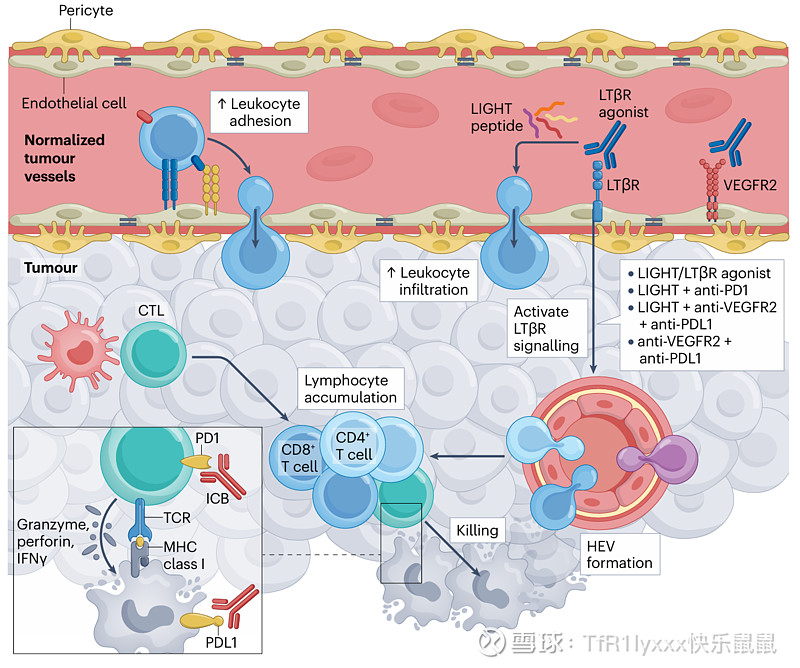
血管正常化还促进了高内皮微静脉(HEVs)的诱导,HEVs 是能够实现淋巴细胞高效运输的特化血管,并可能促进三级淋巴结构(TLSs)的形成。TLSs 的发展可以通过作为抗原呈递和 T 细胞激活的场所来进一步增强局部免疫反应。靶向 LIGHT(一种 LTBR 配体)足以使肿瘤血管正常化,诱导 HEV 发育,增强 CD8+CTL 招募并改善免疫检查点阻断(ICB)的响应。此外,LTBR 激动剂已被证明能在难治性小鼠胶质母细胞瘤模型中诱导 HEV 形成,导致 CTL 激活并使肿瘤对抗血管生成和抗 PDLI 疗法的联合作用敏感。此外,抗 VEGFR2 和抗 PDLI 抗体治疗的组合在各种小鼠癌症模型中通过激活 LTBR 信号通路诱导了 HEV 形成。
图片来源:Kabir AU, Subramanian M, Kwon Y, Choi K. Linking tumour angiogenesis and tumour immunity. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z
血管正常化缓解肿瘤缺氧,增强免疫细胞功能 (Vessel normalization alleviates tumour hypoxia, enhancing immune cell function)
肿瘤缺氧的逆转可以通过正常血管对微环境的充分灌注来实现。确实,使用几种不同的抗血管生成方法使肿瘤血管系统正常化已被证明可以恢复灌注并减少瘤内缺氧。自从最初关于血管减压后肿瘤缺氧解决的报告以来,一些临床前研究报道了在高血管密度肿瘤中肿瘤缺氧的减少。这些包括探索常规抗血管生成靶点(如 VEGF、VEGFR 和 FGF2)的研究,以及包括周细胞、PDGF 和 PDGFR、内皮氧传感器、羟氯喹和致癌负荷减少在内的新的和非传统的靶点。所有这些方法都被认为可以减少肿瘤缺氧,部分原因是血管正常化效应。回顾性和前瞻性研究表明,只有当血管正常化剂的施用与灌注增加和缺氧减少相关时,才能实现最大的治疗益处。来自临床成像引导研究的证据也表明,血管正常化剂的施用改善了缺氧。例如,用西地尼布(cediranib)(一种 VEGF 受体酪氨酸激酶的小分子抑制剂)治疗,通过 MRI 评估,改善了胶质母细胞瘤患者的血液灌注并减少了瘤内缺氧。类似地,使用贝伐珠单抗(一种抗 VEGF 单克隆抗体)的新辅助治疗,通过 18F-misonidazol-PET-PET/计算机断层扫描评估,与乳腺癌患者缺氧减少相关。重要的是要注意,血管正常化后缺氧的减少是一种只能发生在能够耐受“血管正常化窗口期”期间修剪的高密度血管化肿瘤中的现象。需要更好地理解血管正常化窗口及其相关的线索和生物标志物,以便适时施用疗法以获得最大的临床益处。
血管正常化减轻瘤内压力,促进免疫细胞浸润 (Vessel normalization relieves intratumoural pressure, facilitating immune cell infiltration)
间质液压力(IFP)升高是血管异常肿瘤的另一个标志,这是由于渗漏和淋巴系统引流受损造成的。肿瘤内升高的压力不仅促进肿瘤进展,而且作为免疫细胞浸润的屏障,这对于抗肿瘤免疫和免疫疗法的成功至关重要。抗血管生成治疗通过使血管系统正常化,降低 IFP 并恢复 TME 内的正常流体动力学。阻断 VEGFR2 会导致肿瘤进展过程中跨血管壁的静水压力梯度降低。瘤内压力的降低促进了免疫细胞的招募和迁移,使它们能够更深地穿透肿瘤块。当将血管正常化与免疫疗法结合时,这种效果尤其有益,因为降低的压力改善了效应 T 细胞和巨噬细胞浸润肿瘤并与肿瘤细胞接触的能力。此外,通过降低 IFP,血管正常化还增强了在 TME 中积累并损害免疫功能的抗原和免疫抑制因子(如乳酸和腺苷)的排出。
血管正常化与ICB疗法之间的协同作用 (Synergy between vessel normalization and ICB therapies)
癌症免疫治疗,特别是 ICB 治疗,已成为靶向 TME 免疫组成部分的基石。ICB 疗法通过靶向免疫检查点分子(如 PD1 及其配体PDL1)来破坏促肿瘤免疫抑制机制。一个能够带来血管正常化的合理抗血管生成治疗方案可以与 ICB 疗法的免疫刺激效应产生协同作用。确实,临床前研究表明,将 ICB 与抗血管生成治疗相结合可以改善胰腺癌、乳腺癌、肾癌、肺癌、黑色素瘤、卵巢癌、肝细胞癌和结直肠癌模型的肿瘤结局。潜在机制可能包括血管正常化减少瘤内缺氧、增加瘤内效应 T 细胞、减少耗竭 T 细胞的丰度、将 TAM 群体重编程为更具抗肿瘤的M1样表型、促进抗原呈递树突状细胞的成熟以及增强 HEV 的形成等。
联合抗血管生成和 ICB 疗法的有效性提出了一个问题:是否可以根据肿瘤的血管生成和免疫状态来预测治疗结果。为了解决这个问题,我们分析了30种非血液学实体瘤类型的血管生成状态和 T 细胞免疫活性,将患者分为3种不同的血管免疫亚型:(1) 高血管生成伴低抗肿瘤免疫,(2)低血管生成伴高抗肿瘤免疫,以及(3) 中间亚型。有趣的是,血管免疫亚型虽然在肿瘤类型中是保守的,但在不同的实体瘤类型中以不同的比例存在。在所有分析的队列中,在以病理性血管生成活性低和抗肿瘤免疫力高为特征的血管免疫亚型中观察到对ICB的改善反应,这表明TME的这两个组成部分之间存在潜在的生物相互作用,而这种相互作用与个体肿瘤类型的异质性无关。在临床上,这种关系的协同作用最容易在先前已批准抗血管生成疗法或 ICB 疗法的肿瘤类型中进行探索;我们相信这是一种可以加以利用的关系,尽管存在潜在的肿瘤异质性。对我们研究中进行的亚组分析的初步支持了这一观点。肿瘤突变负荷被广泛认为是 ICB 反应的生物标志物。有趣的是,我们通过亚组分析发现,只有当患者属于血管免疫亚型,其病理性血管生成活性较低时,具有高突变负担的患者才能获得生存益处。这表明,血管免疫簇所捕捉的潜在生物学提供了一个统一的框架,用于预测抗肿瘤反应,即使在肿瘤异质性的复杂性中也是如此。我们建议,低血管生成和高抗肿瘤免疫的患者最有可能从 ICB 治疗中受益。相比之下,那些高血管生成和低抗肿瘤免疫的患者可能对先用抗血管生成治疗随后用 ICB 的序贯治疗反应更好。具有中间血管免疫亚型的患者可能对这些治疗策略表现出抵抗,这可以在未来进一步研究。
当前联合抗血管生成疗法和 ICB 的临床试验已在肾细胞癌、HCC 和非小细胞肺癌的治疗中证明了一定的效果。在肾细胞癌中, 靶向 TKI 单药治疗(索拉非尼)一直是标准疗法,抗 PD1 疗法与各种 TKI(如阿昔替尼、乐伐替尼和卡博替尼)的组合与索拉非尼单药治疗相比,在无进展生存期和总生存期方面都显示出显著改善。这在主要临床试验中观察到,如 KEYNOTE-426 (NCT02853331)、CLEAR (NCT02811861) 和 CheckMate 9ER (NCT03141177)。在 HCC 中,IMbrave-150 (NCT03434379) 试验是一个突破,其是第一个成功取得超越索拉非尼生存获益的试验。该试验至关重要,因为它证明了联合疗法对 HCC 的成功,并且是第一个显示免疫疗法作为 HCC 一线治疗选择益处的试验。在其他癌症类型中,包括黑色素瘤、前列腺癌、胃食管癌、乳腺癌和结直肠癌,目前正在进行联合抗血管生成疗法和 ICB 的额外 III 期试验。这些试验旨在进一步评估这种联合策略在将其益处扩展到已显示前景的癌症之外的潜力。
有证据表明,转移性病变对抗pd1单药治疗的耐药性增加。事实上,对病变的事后分析表明,耐药性可能部分归因于CD8+ T细胞在侵袭性肿瘤边缘的浸润不足。尽管这种对单药耐药的确切病理生理机制尚不清楚,但临床数据表明,与单药治疗相比,抗血管生成和ICB联合治疗可改善转移性疾病的反应,这为康方生物AK112提供了很好的指引。
重点来了结论:
最后综上所述,如果把免疫治疗看做一辆载满士兵的火车,那么抗PD-1这样的ICB药物主要是负责识别敌人,不要减速全速前进,而抗VEGF这样的血管正常化药物则主要负责开路,毕竟开的再快没有路也不行,如果路不好也开不太快,毕竟“要想富先修路”,而康方生物AK112无疑是最好的选择,低毒的抗血管,完美的开路者,也就是更好的免疫治疗药物。
最后是一些声明和闲话,本文部分内容出自Kabir AU, Subramanian M, Kwon Y, Choi K. Linking tumour angiogenesis and tumour immunity. Nat Rev Immunol. Published online August 14, 2025. doi:10.1038/s41577-025-01211-z,如有侵权请联系删除。本文仅作学术交流,不构成任何投资建议。风险提示:本文所提到的观点仅代表个人的意见,所涉及标的不作推荐,据此买卖,风险自负。这篇文章难度不小,字数也不少,本来想慢慢写,但是正好赶上了,就一口气都写完了,写的有些匆忙了,希望可以和大家多多交流多多讨论,也希望通过这篇文章我们都能有所收获。谢谢大家了。发出来可能是凌晨了,而且是周末的凌晨,估计看到的人会比较少,大家明天白天再看,也不用熬夜看,如果有能力的,还是推荐看原文的,原文写的真的不错,看完一定有所收获,十分推荐原文。@雪球创作者中心 #雪球星计划# $康方生物(09926)$